钾是植物生长发育所必需的三大营养要素之一,在植物生长发育和叶片衰老过程中,老叶中的K+可经由韧皮部的回流,实现其向根系和新生组织/器官的再分配过程。目前对这一过程的分子机制的认识还不尽明晰。已知在模式植物拟南芥中,AtAKT2钾离子通道具有介导韧皮部K+回流的部分功能。在功能方面,AtAKT2兼具K+吸收和外排活性,即弱电压依赖型双向整流性质;其生理意义在很大程度上体现在其K+外排活性对韧皮部细胞极化状态的平衡方面,因而有助于地上部光合同化产物蔗糖经韧皮部途径的向下运输过程。目前所知的AKT2通道,尤其是在双子叶植物中,大多具有这些特征。
我室苏彦华研究员课题组通过对水稻基因组中的同源钾通道OsAKT2的分析发现,与拟南芥等植物中的典型AKT2通道基因相比,OsAKT2的膜电位感应区(S4跨膜区)中编码精氨酸R的碱基发生了由AGA到AAG的序列顺序置换,而导致该氨基酸变异为赖氨酸K。通过蛙卵双电极电压钳电生理技术进行了系统的功能解析,证明水稻OsAKT2明显不同于典型的双向整流型AKT2钾通道,其主要介导K+的吸收(内流)而缺失了该型通道的标志性特征—K+外排(外流)活性,且OsAKT2通道的行为表现出强烈的电压依赖性。通过定点突变技术消除OsAKT2中的上述氨基酸变异(即K191R点突变),通道随即恢复了与典型AKT2类通道相同的双向整流性;而将拟南芥AtAKT2的对应氨基酸R改变为K,则其原有的外排活性丧失。由此可见水稻OsAKT2功能的改变来源于其K191氨基酸编码序列的物种变异—K191特异性氨基酸的变异锁住了OsAKT2的K+外排活性(图1)。同时,OsAKT2所缺失的K+外排活性可被OsCBL1/OsCIPK23复合磷酸化过程部分激活(图2)。该文揭示了水稻OsAKT2钾通道功能的这一结构性锁定—磷酸化解锁机制。进一步利用AKT2功能缺陷型拟南芥akt2突变体作为工具,证实了水稻OsAKT2通过该位点的自然变异而强化其韧皮部K+回流的功能(图3)。
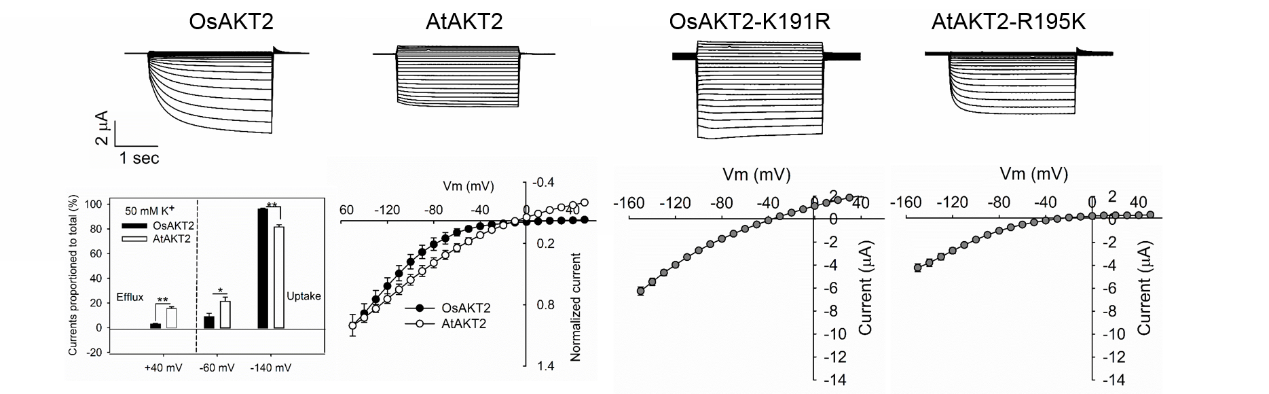
图1. 特异性K191氨基酸的存在,将OsAKT2通道的活性锁定在仅介导K+吸收的状态
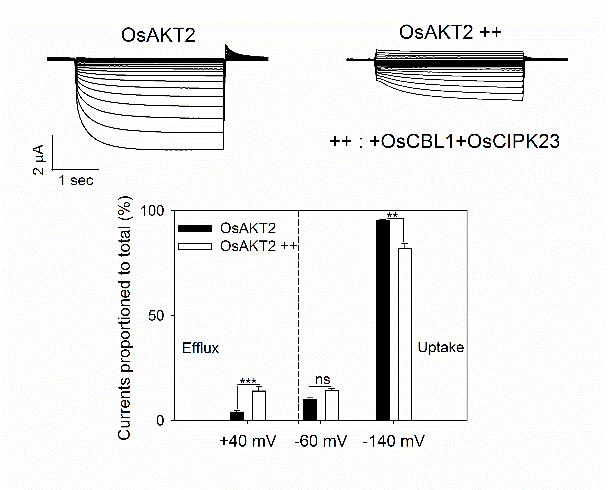
图2. OsAKT2的K+外排活性可被OsCBL1/OsCIPK23磷酸化机制部分解锁
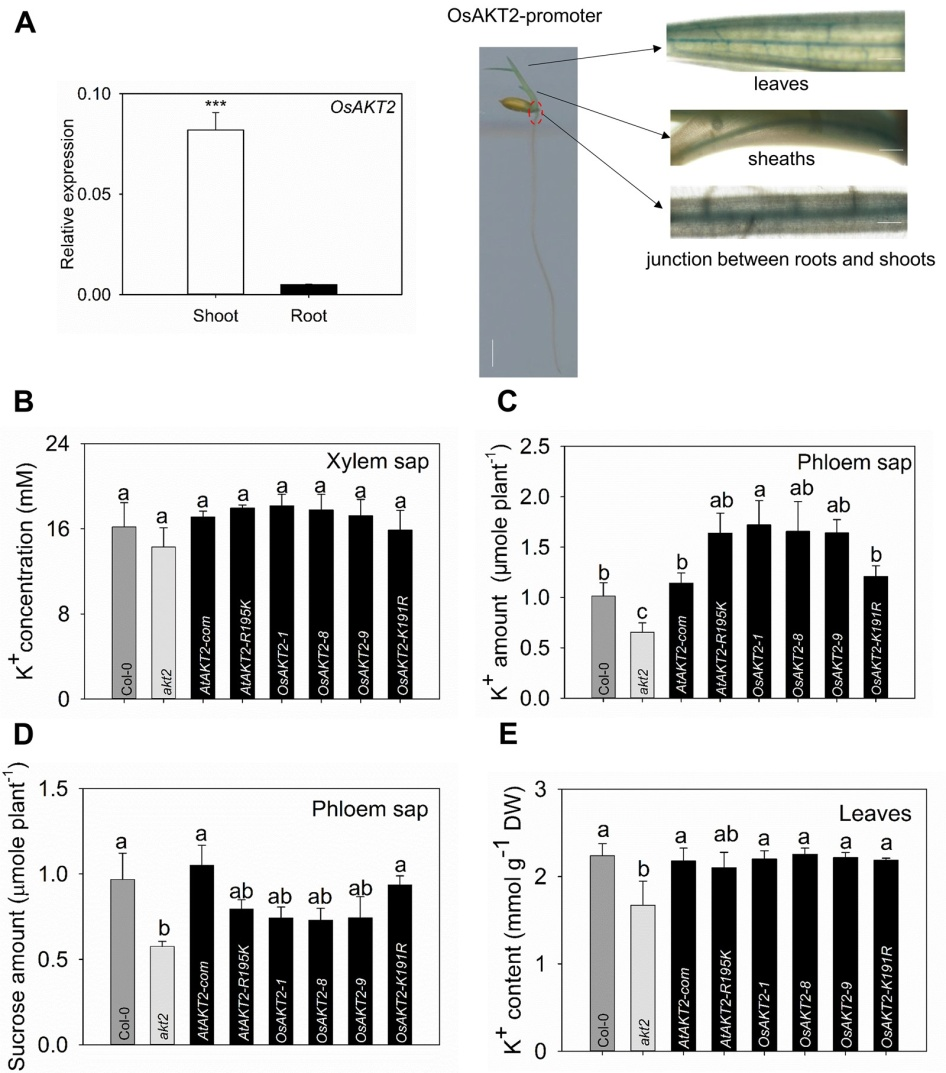
图3. OsAKT2介导地上韧皮部K+的装载和再循环
上述研究成果发表在Plant Physiology上,苏彦华研究员为文章通讯作者,博士生黄亚楠为文章第一作者。该研究得到国家重点研发专项和国家自然科学基金的资助。
原文链接:
https://academic.oup.com/plphys/advance-article/doi/10.1093/plphys/kiab462/6380557





